Introduction to the Arthropoda
. . . the REAL rulers of the Earth. . .

By nearly any measure, the most successful animals on the planet are the arthropods. They have conquered land, sea and air, and make up over three-fourths of all currently known living and fossil organisms, or over one million species in all. Since many arthropod species remain undocumented or undiscovered, especially in tropical rain forests, the true number of living arthropod species is probably in the tens of millions. One recent conservative estimate puts the number of arthropod species in tropical forests at
Arthropods range in distribution from the deep sea to mountain peaks, in size from the king crab with its 12-foot armspan to microscopic insects and crustaceans, and in taste from chocolate covered ants to crawfish jambalaya and lobster Newburg. Despite this unbelievable diversity, the basic body plan of arthropods is fairly constant. Arthropods have a stiff cuticle made largely of chitin and proteins, forming an exoskeleton that may or may not be further stiffened with calcium carbonate. They have segmented bodies and show various patterns of segment fusion (tagmosis) to form integrated units (heads, abdomens, and so on). The phylum takes its name from its distinctive jointed appendages, which may be modified in a number of ways to form antennae, mouthparts, and reproductive organs
Characteristics
· Makes up 3/4's of all animal species -total number of arthropod species is MORE than all other species combined
· Includes insects, spiders, & crustaceans
· Arthropod means "jointed foot" - all arthropods have jointed appendages
· Segmented body
- Exoskeleton for protection & support
- Exoskeleton is shed during molting
· Open circulatory system
· Compound eyes (has many individual units)
· Excretory structures called Malpighian tubules
- Wings in many groups
- Respiration using spiracles and trachae
Segmentation
More obvious in larval forms, adults have fused segments -----> Head | Thorax | Abdomen
Some have a fused head and thorax -- the cephalothorax
Taxonomy
Arthropods have traditionally been divided into four classes: Trilobita, Chelicerata, Crustacea, and Uniramia. Most living and post-Cambrian fossil arthropods fall into one of these four classes. However, a number of arthropod fossils from the Cambrian Period have been described which do not obviously belong in any of these classes. Many of these arthropods have been found in the famous Middle Cambrian Burgess Shale, in the Canadian Rocky Mountains of British Columbia. Other "weird wonder" arthropods have been found in
Debate as to which living phyla are the closest relatives of arthropods continues to rage. Arthropods were once considered to be close relatives of annelids, or segmented worms, but this is no longer universally accepted (Eernisse et al. 1992), and so far, no consensus has emerged from molecular studies. However, on both morphological and molecular grounds it seems that two groups that are sometimes given phylum rank, the Onychophora and the Tardigrada, are the closest living relatives of the arthropods. Some Vendian and Cambrian organisms that are not arthropods may have been even closer to living arthropods, although finding their precise position is difficult. For the purposes of this exhibit, these are classified as the "vendiamorphs" and the "anomalocaridiids." The cladogram used in this virtual exhibit is a simplification of that in Wills et al. (1994), with some additions based on Eernisse et al. (1992) and Waggoner (1996). It should be emphasized that arthropod phylogeny is often hotly debated, and a number of alternative hypotheses have been proposed.
Systematics of the Arthropoda

Arthropods have traditionally been divided into four classes: Trilobita, Chelicerata, Crustacea, and Uniramia. Most living and post-Cambrian fossil arthropods fall into one of these four classes. However, a number of arthropod fossils from the Cambrian Period have been described which do not obviously belong in any of these classes. Many of these arthropods have been found in the famous Middle Cambrian Burgess Shale, in the Canadian Rocky Mountains of British Columbia. Other "weird wonder" arthropods have been found in
Debate as to which living phyla are the closest relatives of arthropods continues to rage. Arthropods were once considered to be close relatives of annelids, or segmented worms, but this is no longer universally accepted (Eernisse et al. 1992), and so far, no consensus has emerged from molecular studies. However, on both morphological and molecular grounds it seems that two groups that are sometimes given phylum rank, the Onychophora and the Tardigrada, are the closest living relatives of the arthropods. Some Vendian and Cambrian organisms that are not arthropods may have been even closer to living arthropods, although finding their precise position is difficult. For the purposes of this exhibit, these are classified as the "vendiamorphs" and the "anomalocaridiids." The cladogram used in this virtual exhibit is a simplification of that in Wills et al. (1994), with some additions based on Eernisse et al. (1992) and Waggoner (1996). It should be emphasized that arthropod phylogeny is often hotly debated, and a number of alternative hypotheses have been proposed.
 Introduction to the Spriggina
Introduction to the Spriggina
The striking Vendian fossil Spriggina (shown here) and its close relative Marywadea make up the Sprigginida, a clade of soft-bodied organisms that are restricted to the Precambrian. Spriggina is known largely from the Ediacara Hills of
Spriggina was described as an annelid (segmented worm), but it now appears to be related to the arthropods, although Spriggina had no hard parts, and it is unclear exactly what kind of appendages it had.
Introduction to the Pycnogonida

Pycnogonids, or "sea spiders", are among the most bizarre-looking arthropods. Another name sometimes used for them, Pantopoda, means "all legs" and describes them perfectly. Pycnogonids have extremely reduced bodies in which the abdomen has almost disappeared, while the legs are long and clawed. The head has a long proboscis with an unusual terminal mouth and several simple eyes on a central tubercle. The head also bears a pair of claws and a pair of ovigers on which the eggs are carried. All in all, it can be hard to tell just which end of a pycnogonid is the head; in this picture the head is to the right (we think) and the proboscis has been bent under the body.
Pycnogonids feed on soft-bodied invertebrates, in particular cnidarians, sucking at them with their probosces, and larval pycnogonids often live as parasites within cnidarian tissues. The intestine of pycnogonids has extremely long diverticulae (blind pouches) that extend to the ends of the legs.
Pycnogonids have almost no fossil record. Three genera have been found in the Devonian, in the Hunsruck Slate of western
Introduction to the Onychophora
"Velvet worms" with an ancient history indeed

The average resident of the Northern Hemisphere is probably not familiar with the Onychophora; they are restricted to forest regions of South America , Africa , the Caribbean , and Oceania . Shy creatures, able to hide in incredibly tight crevices, these "velvet worms" (about ninety living species known) are rarely seen even in their natural habitat. Yet onychophorans are of great interest to biologists, because they seem to be related to arthropods, and give us an idea of what the ancestors of the arthropods may have been like. Although they are rare as fossils, a number that have been found from the Cambrian period. These fossils show that abundant marine relatives of the Onychophora flourished in the seas 520 million years ago.
Introduction to the Trilobita
Gone, but not forgotten. . .

Among famous groups of extinct organisms, trilobites probably rank second only to the dinosaurs in fame. Their fossils are instantly recognizable and often strikingly beautiful. In fact, prehistoric people liked them as much as we do. At Arcy-sur-Cure in
Although the last few trilobites went extinct about 245 million years ago, they are one of the best-known and most-studied groups of fossil arthropods.
Trilobita:Systematics

As traditionally defined, the true Trilobita are divided into two main groups, the Agnostida and the Polymerida. Agnostids have sometimes been considered non-trilobites -- even crustaceans, by some -- but recent analyses (e.g. Fortey and Theron 1994) support the inclusion of the Agnostida in the Trilobita.
A third group, the Nektaspida, was not identified until the 1980s; its members include soft-bodied arthropods from the Burgess Shale such as Naraoia. Some, such as Gould (1991), have classified the Nektaspida as true trilobites; others (e.g. Simonetta and
Many other Cambrian arthropods, especially those from the world-famous Burgess Shale, were once considered probable trilobite relatives, or "trilobitomorphs." While many of these problematic arthropods are no longer thought to be related to the true trilobites, recent studies have shown that a number of them are closer to the trilobites than to any other arthropods. The term "Trilobitomorpha" is used here in this more restricted sense. Exactly how these arthropods are related to the trilobites is not yet perfectly clear, but the non-trilobite "trilobitomorphs" probably compose several clades. One of the clades that probably fits in here is called the Emeraldellida. Yet another group of arthropods, the aglaspids, have traditionally been grouped with the chelicerates, but may be closer to trilobites. Some authors also group the emeraldellids with the cheliceramorphs as well (e.g. Simonetta and
Introduction to the Polymerida
 The Polymerida, which make up 95% of all trilobites, are the trilobites that most people are familiar with. Unlike the agnostids, polymerid trilobites typically have more than two or three thoracic segments, and the pygidium is usually smaller than the cephalon. Unlike the blind agnostids, most polymerid trilobites had remarkably well-developed compound eyes. (A few, like Ductina, were blind, but this was a secondary evolutionary development.) Polymerid trilobites appear in the Early Cambrian Period, and the last ones went extinct at the end of the Permian.
The Polymerida, which make up 95% of all trilobites, are the trilobites that most people are familiar with. Unlike the agnostids, polymerid trilobites typically have more than two or three thoracic segments, and the pygidium is usually smaller than the cephalon. Unlike the blind agnostids, most polymerid trilobites had remarkably well-developed compound eyes. (A few, like Ductina, were blind, but this was a secondary evolutionary development.) Polymerid trilobites appear in the Early Cambrian Period, and the last ones went extinct at the end of the Permian.
There is currently no consensus as to how the major groups of polymerid trilobites are related to each other. In fact, it is possible that the group is paraphyletic; the last common ancestor of all polymerid trilobites may also have been ancestral to the agnostids as well.
The text below presents the polymerid trilobites as they are traditionally classified, with no evolutionary relationships indicated. For the moment, click on any of these in-line pictures for a full picture, to get an idea of trilobite diversity.
Order Redlichiida | |||||
| | | | Suborder Redlichiina |
| Bergeroniellus spinosus Early Cambrian |
| | | | |
| Paradoxites sp. Cambrian |
| | | | Suborder Olenellina |
| Nevadella sp. Early Cambrian |
Order Corynexochiida | |||||
| | | | |
| Olenoides serratus Middle Cambrian |
| | | | |
| Ogygiocarella debuchii Middle Ordovician |
Order Ptychopariida | |||||
| | | | Suborder Ptychopariina |
| Elrathia kingi Middle Cambrian |
| | | | Suborder Asaphina |
| Megistaspidella sp. Lower Ordovician |
| | | | |
| Homotelus sp. Ordovician |
| | | | Suborder Illaenina | | |
| | | | Suborder Harpina | | |
| | | | Suborder Trinucleina | | |
Order Phacopida | |||||
| | | | Suborder Cheirurina |
| Gabriceraurus sp. Middle Ordovician |
| | | | |
| Pliomera fischeri Ordovician |
| | | | Suborder Phacopina |
| Dalmanites limuloides Silurian |
| | | | |
| Ductina vietnami Devonian |
| | | | |
| Phacops rana Devonian |
| | | | Suborder Calymenina |
| Flexicalymene niagarensis Silurian |
Order Lichiida | |||||
Order Odontopleurida | |||||





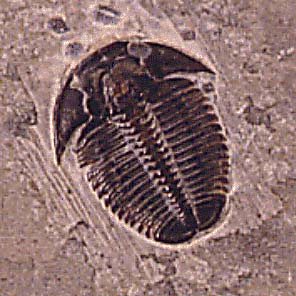





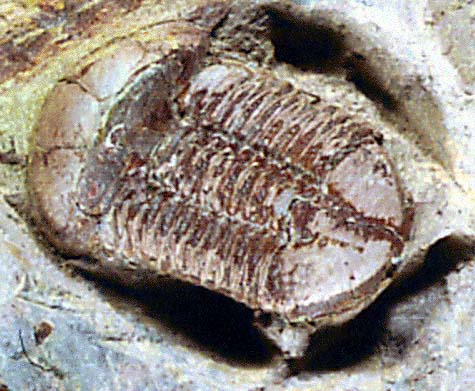


Introduction to the Agnostida


Agnostid trilobites were rather unusual trilobites: it is thought by some paleontologists that they may have been planktonic. They are small, often lack eyes, and have a pygidium the same size as the cephalon; it can be difficult to tell which end of an agnostid is the front! Agnostids have only two or three segments between the cephalon and pygidium.
Agnostids appeared in the Lower Cambrian and are common in some Cambrian rocks; they became rare in the Ordovician and went extinct at the end of the Ordovician. They are useful for biostratigraphic correlation of Cambrian rocks.
There are two subgroups of agnostids. The Agnostina is represented at the top of the page, by specimens probably in the genus Ptychagnostus, from the Middle Cambrian of Utah. These trilobites are only about 8 mm (about 1/3 inch) long. Members of the Agnostina have only two segments between the cephalon and pygidium, and never have cephalic sutures. Below is a picture of Pagetiellus, from the Lower Cambrian of the
Introduction to the Cheliceramorpha

The Cheliceramorpha includes spiders and scorpions, mites and ticks, horseshoe crabs, daddy-longlegs, and extinct "sea-scorpions", to name a few. It is the second most prominent order of terrestrial arthropods, after the uniramians. Most of its marine representatives are extinct, but were prominent in the Paleozoic and included some fearsome predators.
Cheliceramorpha: Part 1

There are a number of Cambrian fossils that indicate that the Cheliceramorpha was once much more diverse than it is now; these fossils do not fit into any of the above clades. For purposes of clarity, we have left them out of this cladogram, especially since their precise systematic position is debatable and subject to change (specialists should consult Wills et al., 1994).
Traditionally, the Xiphosurida (horseshoe crabs) and Eurypterida (sea scorpions) have been grouped together in a taxon called the Merostomata, while the true scorpions have been classified in the Arachnida along with the spiders, mites, and others. All three of these taxa together form a "crown group", the Chelicerata; the Cheliceramorpha would include the Chelicerata plus all extinct lineages that are closer to it than to any other clade. The Merostomata, however, is probably a paraphyletic group. In fact, there is some evidence to suggest that the terrestrial scorpions are closely related to the eurypterids or sea scorpions (Meglitsch and Schram, 1991). Some recent findings suggest that the Aglaspida, a group whose affinities have always been problematic, may in fact be closer to the trilobites than to the other chelicerates.
Xiphosura
Horseshoe crabs
Unlike all other living chelicerates, horseshoe crabs have compound eyes. However, the eyes appear smooth because the many subunits of the eyes, or ommatidia, are covered by a single thick cornea. There are also two simple eyes, or ocelli, in the center of the head.
Like all chelicerates, Xiphosurans have a body divided into two main subdivisions, the prosoma ("head") and opisthosoma. Six pairs of legs are present on the prosoma (seven in a few fossil species). These legs bear claws except for the last pair. The appendages on the opisthosoma are modified into a series of overlapping plates that are used as gills for respiration. Behind the opisthosoma is a telson, which is elongated into a long spine, and which gives the group its name (xiphos is Greek for "sword," and uros is Greek for "tail").
|
|
|


Horseshoe Crabs : On the left, a ventral (underside) view of a modern horseshoe crab Limulus polyphemus, showing the many legs and gills. On the right, a dorsal (back) view, distinctly showing the two body divisions and the telson. Notice that each of the two body sections is expanded outward, providing a protective armor which shields the legs and gills.
 Xiphosurans have existed since the Silurian with relatively little morphological change. The main trends in the evolution of the xiphosurans have been an overall increase in size, a loss of segmentation on the opisthosoma, and a restriction to marine habitats. Whereas all living xiphosurans are marine, it seems that some late Paleozoic and Mesozoic forms inhabited brackish or even fresh water. Below at right, is a picture of Palaeolimulus, a Paleozoic xiphosuran from Mazon Creek,
Xiphosurans have existed since the Silurian with relatively little morphological change. The main trends in the evolution of the xiphosurans have been an overall increase in size, a loss of segmentation on the opisthosoma, and a restriction to marine habitats. Whereas all living xiphosurans are marine, it seems that some late Paleozoic and Mesozoic forms inhabited brackish or even fresh water. Below at right, is a picture of Palaeolimulus, a Paleozoic xiphosuran from Mazon Creek,
The Xiphosura are not a diverse group in the oceans of today. One species, Limulus polyphemus (shown above), lives along the
Introduction to the Eurypterida
 There are more than 300 species of sea scorpions known. These have been classified in more than 60 genera and about 20 families. Most major groups can be recognized by the structure of the legs and chelicerae, but many genera and species may only be recognized by structures on the ventral (belly) side of the animal. For fossils in which only the dorsal (back) side is visible, this means we cannot always identify them to species. The fossil at right is a remarkable exception; two fossils of Eurypterus were fossilized side-by-side showing both the dorsal and ventral aspect.
There are more than 300 species of sea scorpions known. These have been classified in more than 60 genera and about 20 families. Most major groups can be recognized by the structure of the legs and chelicerae, but many genera and species may only be recognized by structures on the ventral (belly) side of the animal. For fossils in which only the dorsal (back) side is visible, this means we cannot always identify them to species. The fossil at right is a remarkable exception; two fossils of Eurypterus were fossilized side-by-side showing both the dorsal and ventral aspect.
The most recent complete revision of eurypterid classification was done by V.P. Tollerton, Jr., in 1989. He recognized a dozen superfamilies based on the morphology of the legs — whether they were spiny, similar in appearance, and whether the sixth pair of appendages were modified as paddles. He recognized two suborders, the Pterygotina with greatly enlarged pincer-like chelicerae and simple walking legs, and the Eurypterina in which the chelicerae are small and the forward legs are usually spiny.
Unfortunately, there are no published cladistic studies on the Eurypterida, and only a single unpublished thesis attempting to produce a phylogeny of all the taxa. It therefore remains uncertain which groups and what characters are truly primitive among sea scorpions.
Eurypterids are chelicerate arthropods, and until recently were put into the Class Merostomata with the Xiphosura (horseshoe crabs). It is now believed that the Eurypterida is the sister group to the Arachnida, making the sea scorpions closer relatives of the scorpions and spiders than to horseshoe crabs.
Jason Dunlop of the

The prosoma of eurypterids is both the head and forebody of the animal. The dorsal (top) side of the prosoma is a single plate from which protrude two compound eyes at the sides, and two smaller ocelli (simple eyes) from the top central portion. The eurypterid shown above (Pterygotus) has unusually large compound eyes. The ventral (belly) side of the prosoma may be a single shield-like plate or many small plates. The mouth is underneath in the center, where the legs are attached.
There are six pairs of appendages attached to the prosoma. The first pair are the chelicerae, which are homologous to the fangs of spiders. The chelicerae are not homologous to the pincers of scorpions, despite their resemblance in the diagram above; scorpion pincers are the second appendages, not the first. In most sea scorpions, the chelicerae are rather small; only the Pterygotina have chelicerae as large as those shown above. The next four pairs of appendages are the walking legs. These may have spines, and may come all in a single shape (as above), or may be of several sizes and shapes as in the genus Mixopterus. The final pair of appendages may also be legs in some species, but usually are modified into paddles.
The opisthosoma always has twelve segments called tergites. The first six are usually broader than the last six, and so are given the special name mesosoma, meaning "middle body". The mesosomal tergites are the segments that bear the five pairs of gills, which are covered by protective plates. The first pair of gills/plates also contain the reproductive organs. The remaining six segments form a narrower "tail" that may be flattened or tube-shaped. At the end of the tail is the telson. This final segment may be a spike, a paddle, or a set of pincers.
Far less is known about the internal structure of sea scorpions. There are a few fossils in which part of the intestine is preserved, and this ends in an opening between the final tergite and the telson, on the underside of the tail.
Cheliceramorpha:Part II
The Arachnida

The Arachnida include the terrestrial chelicerates that everyone is familiar with, and that nearly everyone would rather not be too familiar with: spiders (Araneae), ticks and mites (Acari), and scorpions (Scorpiones). Arachnids also include a number of less familiar taxa: Opiliones (harvestmen or daddy-longlegs); Thelyphonida (whip-scorpions); Pseudoscorpiones (false scorpions); and many others. Most are predators, and some are venomous. All are terrestrial, except for some mites and spiders that have become secondarily aquatic.
Araneae
Spiders


The Araneae are the true spiders. Unlike the fossil spider-like trigonotarbids and their allies, almost all spiders have only five segments in the abdomen, and these are generally fused with no external trace of segmentation -- the earliest spiders had as many as twelve segments in their abdomen.
The last two abdominal segments are specially modified into spinnerets which secrete the silk threads for which spiders have become well known. There are one to four pairs of spinnerets present, even on those spiders which do not spin webs. The silk has many other functions, such as in sperm transfer, encasing the eggs, and building nests or burrows. Those spiders which do use their silk for webs often produce complex and intricate patterns, but complex or not, the web's function is the capture of prey.
Other spiders which do not spin webs will stalk or ambush their prey. Wolf spiders, tarantulas, and jumping spiders are of this sort. Some species are brightly colored, and hide within flowers where they are camouflaged, waiting to pounce on visiting insects. A very few of these are large enough to capture small birds. These spiders rely on their amazing speed and paralyzing poison to subdue their captures.
The front pair of appendages, the chelicerae, are the ones which contain the poison glands. The second pair, the pedipalps, are small, and are used by the male during mating. The head bears four pairs of eyes, the arrangement of which can be very useful for the identification of the several different kinds of spider. Most spiders breathe through tracheae; some, like the wolf spiders, have both book lungs and tracheae.
As with other arachnids, there is an abundance of fossils in the mid-Paleozoic and the later part of the Cenozoic. However, there are no known spiders from the Mesozoic, a gap in the record of 200 million years.
Trigonotarbida and their Allies

A number of Carboniferous arachnids somewhat resembled spiders, and in fact appeared at about the same time as the first true spiders. Like spiders, they were terrestrial and respired through book lungs, and walked on eight legs. However, they were not true spiders, nor necessarily ancestral to them, but represented independent offshoots of the Arachnida. Trigonotarbids are currently the oldest known land arthropods. Most of these fossils are found in the Upper Carboniferous (Pennsylvanian), though they have also been found in the Ludlow Lane fauna of England (Pridolian, late Silurian), the Alken an der Mosel fauna of Germany (Emsian, early Devonian) the Rhynie Chert of Scotland (Siegenian, early Devonian) and the Gilboa, New York fauna from the Panther Mountain formation (Givetian, middle Devonian). Not much is known about their relationships; we present them on one page primarily for convenience.
Eophrynus prestvichi, shown above as illustrated in Karl Zittel's 19th-century textbook of paleontology, is from the Upper Carboniferous of England. It belongs to one of the better-known Carboniferous arachnid groups, the Trigonotarbida, which may be most closely related to a group of soft-bodied, small arachnids known as the Palpigrada. Unlike true spiders, Eophrynus has a broad junction between prosoma ("head") and opisthosoma "abdomen"). Another trigonotarbid, Pleophrynus, from the Mazon Creek deposits of
Architarbids were another Late Carboniferous group of arachnids. Orthotarbus, also from Mazon Creek, is one example of an architarbid. Note that the abdomen has ten segments; the first five are abbreviated and thickened, while the last five are broad and indistinctly demarcated. The specimen is also from the Illinois State Museum.
Acari
Mites and Ticks


The Acari include ticks, mites, and their kin. The black-legged tick, or deer tick, Ixodes scapularis, shown here is a relatively large member of the Acari, about 5 mm long. Many other members of the Acari are microscopic, like this unidentified mite photographed with UCMP's Environmental Scanning Electron Microscope.
With small size has come morphological change: the segmentation of the opisthosoma and the distinction between prosoma and opisthosoma has been almost completely lost, as have the heart and eyes in many species. The chelicerae (first appendages) may be clawed mouthparts or may be modified for piercing, as in ticks. Some mites have even reduced or lost one, two, or three pairs of legs.
Mites and ticks which feed on vertebrate hair or blood often carry disease organisms, such as spirochete bacteria, responsible for relapsing fever and Lyme disease. Others are rather unpleasant parasites themselves, such as ticks, chiggers, and the skin mites that cause mange and scabies. Yet most mites are free-living, found in great abundance in soils, plant litter, and even in water. Those that parasitize agricultural insect pests may be beneficial. In all there are about 30,000 species of Acari known, and that is probably only a fraction of the actual number of species.
Mites were among the first land arthropods; the oldest known so far are Late Devonian. However, because of their small size, the Acari are uncommon in the fossil record. A few are known from the Carboniferous, and more from Tertiary amber.
 Scorpiones
Scorpiones
Scorpions are the oldest arachnids for which fossils are known, and they were the first arachnid fossils to be found in Paleozoic strata. The Silurian scorpions appear to have lived in the water, since their fossils have gills, but by the Carboniferous scorpions with such features are no longer found -- fossils from the Pennsylvanian age Mazon Creek beds have book lungs covered by protective plates, and so were probably land-dwellers. The best scorpion fossils come from the Devonian and the Oligocene; there is a severe lack of fossils known from the intervening period.
These earliest scorpiones are considered to be Protoscorpions, since they possess many traits which are plesiomorphic for scorpions. For example, in all scorpions the thick front portion on the abdomen is made up of seven segments, but the number of sternite plates which cover this region varies among the earliest fossils, while all living species have five. All scorpions have an additional five segments after the initial seven, ending in a sharp sting. This sting contains a pair of poison glands which can paralyze prey, usually insects or small rodents, or may deliver a painful sting to incautious persons. Most scorpion stings are merely painful, leading to swelling in the immediate region of the sting, but some scorpions of northern
Besides their unusually long and dangerous tails, scorpions also differ from other arachnids in having large pedipalps. These are the second pair of appendages on the body, and are usually rather inconspicuous in arachnids, but in scorpions, they are large and powerful pincers which may be used to grasp and subdue prey. Scorpions may also have more eyes than other arachnids, some species possessing as many as six pairs, though most do not have this many. They have three joints in their chelicerae, or the first pair of appendages, located next to the mouth.
Most scorpions are nocturnal, hiding under rocks, in crevices, or within burrows during the day, and coming out after sunset. Because of this, and because of their painful stings, it can be dangerous to travel at night in scorpion territory without shoes, even inside homes. (I have nearly stepped on a scorpion in my parents' home near
Crustaceamorpha:Systematics
| | |||||||||||||||||||||||||||||||||||||||


| Crustaceans, like other Arthropods, have an exoskeleton made of chitin and proteins that protects them from drying out and provides a hard framework to which their muscles attach. Because of their hard exoskeleton, to increase in size they must molt or shed their exoskeleton and produce a new one (see box at left). Between the time that they shed the old exoskeleton and produce a new one is a very vulnerable time for crustaceans because they are easy prey for many predators. Crustaceans have an open circulatory system meaning that all their blood is not contained within vessels, instead, blood is drawn in to the heart through holes called ostia, then pumped out again to circulate through the tissues and return again to the heart.
| ||||||||||||||||||||||||
| Crustaceans are usually segmented with one pair of appendages per segment. Segments are arranged into distinct tagmata (regions that are speciallized for specific purposes) such as the cephalon (head, cephalic = adjective), thorax (thoracic = adjective), and abdomen. In some taxa the thorax and abdomen are not distinguished from each other and are then called the trunk. Within a region, fusion of segments is common, particularly in the cephalon where it is seldom possible to differentiate each segment. The fusion of the cephalic segments is most obvious when a cephalic shield or carapace is present. A cephalic shield is a fusion of the dorsal (normally the uppermost part, in humans it is the back) portion of the cephalic segments. A carapace, or cephalothorax, is similar to a cephalic shield but is fused further around to the ventral (underside) portion. The table below summarizes the general morphology of the different crustacean groups.
| |||||||||||||||||||||||||
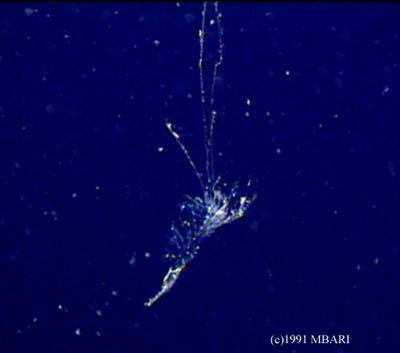

The specialized crustaceans living today are quite different from what ancestoral crustaceans are thought to have been like. They were thought to have consisted of many similar segments, each with a pair of biramous appendages that were also similar to each other. All the variations in morphology seen today in the crustacean groups arose from ancestors with this basic body shape. The Remipedia most closely resemble this ancestor, though they do have several differences and specializations.
Introduction to Maxillopoda
The little ones!
Maxillopods are generally small animals with barnacles being the exception to this rule. They commonly have shortened bodies, with a reduced abdomen that typically lacks appendages. It has been suggested that the ancestor common to all maxillopods developed sexual maturity before it had completely metamorphosed into the adult form. If this hypothesis is correct, it would explain the small size, shortened body plan and reduced number of appendages typically found in Maxillopoda.
Ostracods, copepods and barnacles are the most abundant and diverse groups within the Maxillopoda. Ostracods and barnacles have substantial fossil records. Mystacocarida are tiny worm-like crustaceans. Branchiura and Tantulocarida are both tiny ectoparasitic crustacean groups.
Choose from the cladogram below to begin exploring theis diverse group of crustaceans. As are many groups, the classification of the Maxillopoda is controversial so here we give a generally accepted phylogeny that is subject to change as the groups are studied further.
Crusteceamorpha:Part II
Maxillopoda


As are many groups, the classification of the Maxillopoda is controversial so here we give a generally accepted phylogeny that is subject to change as the groups are studied further.
Introduction to the Branchiura
Fish Lice!


Left: dorsal view of branchiuran Argulus japonicus
Right: ventral view of branchiuran Argulus japonicus, courtesy of David Allard (TAMU)
The Branchiura are a parasitic crustacean group of uncertain relationship within the Maxillopoda. They are thought to be an ancient group but there are no known fossils. There are approximately 130 species described currently, all of which are ectoparasites (attach to the outside of the host) on fish. A few exceptions have been found attached to amphibians.
The branchiurans' very flat, oval body is covered almost completely by their broad carapace with unusual, prominent compound eyes. Branchiuran mouthparts and antennae are modified for their parasitic lifestyle and form a proboscis with hooks, spines, and suckers. They are capable of leaving their host for up to three weeks. Branchiurans leave their hosts for several reasons: to find a different host, to mate, and to lay eggs. When they locate a new host they grab a hold, scurry toward the head, and attach behind the operculum (the flap that covers a fish's gills) or a fin. Attaching in these slightly protected areas reduces the likelihood they will be brushed or washed off of the host. Once attached to the host they either pierce the hosts skin and suck blood and other internal fluids, or they feed on mucus and skin sloughed off by the host. Sever infestations can devastate and aquarium. They can move very fast which is rather disconcerting when watching a swarm in your aquarium.
Introduction to the Ostracoda
Seed or Mussel Shrimp
 The Ostracoda are one of the most successful crustacean groups with approximately 8000 living species. Ostracods are generally small, ranging in length from 0.1 to 32 mm (that's smaller than a poppy seed to the size of a meatball). As indicated by its name, Gigantocypris a planktonic ostracod, is by far the largest member of this group reaching up to 32 mm. Gigantocypris's pelagic life style (continuously swimming in the open water) sets it apart from many other ostracods as well. Most other ostracods are found crawling on or burrowing into the sediments at the bottom of the ocean or lakes. A few species, for example Mesocypris sp., are also found crawling around in moist terrestrial habitats such as mosses. In these habitats, they feed on dead organic material, suspended organic particles, microscopic plants, or they are predators.
The Ostracoda are one of the most successful crustacean groups with approximately 8000 living species. Ostracods are generally small, ranging in length from 0.1 to 32 mm (that's smaller than a poppy seed to the size of a meatball). As indicated by its name, Gigantocypris a planktonic ostracod, is by far the largest member of this group reaching up to 32 mm. Gigantocypris's pelagic life style (continuously swimming in the open water) sets it apart from many other ostracods as well. Most other ostracods are found crawling on or burrowing into the sediments at the bottom of the ocean or lakes. A few species, for example Mesocypris sp., are also found crawling around in moist terrestrial habitats such as mosses. In these habitats, they feed on dead organic material, suspended organic particles, microscopic plants, or they are predators.
Ostracods consist of little more than a head. They have the typical five pairs of appendages on their head but only 1-3 pairs of appendages on the rest of the body. Their bivalved carapace may cause you to mistake them for tiny clams or mussels, thus the common name of "mussel shrimp". The two part, hinged carapace encloses the entire body, similar to the branchiopod Conchostraca. However, their appendages distinguish them from the conchostracans. Another feature that differentiates these groups is a lack of growth rings on the carapace. Ostracods shed the carapace with each molt, whereas the conchostracans simply add material to the carapace as they grow.
The ostracods have the most complete fossil record of any of the crustaceans.
Introduction to the Cirripedia
Barnacles and their Relatives
| Gooseneck barnacle from the |
|
|

There are approximately 1000 species belonging to four groups within the Cirripedia. They are the acorn and gooseneck barnacles (Thoracica), the tiny Acrothoracica, and two parasitic groups, the Rhizocephala and Ascothoracica. Charles Darwin spent much time studying barnacles and wrote four important monographs in the 1850's on living and fossil cirriped groups. These works are still referenced today because of his careful observations and the meticulus illustrations.
Tiny Acrothoracica burrow into calcareous substrates such as mollusk shells or coral skeleton. Because they burrow into hard substrates, they only have to build their own calcareous plates to cover the opening of their burrow. Acorn barnacles on the other hand, secrete calcareous plates to completely cover their bodies. These plates protect the animal from many predators and from drying out when they are exposed to the air during low tides. Acorn barnacles (Thoracica) are the rough white barnacles seen on rocks in the intertidal all over the world. Members of the Thoracica are found many other places besides intertidal rocks. A giant acorn barnacle the size of a softball is found in the depths of the Southern Ocean off of
Ascothoracians are sucking ectoparasites on anthozoans (anemones and kin) and echinoderms whereas rhizocephalans are endoparasites on decapods (crabs and shrimp). Members of these parasitic groups are seldom seen by anyone other than specialists. However, if you were to examine a fresh catch of shrimp you would likely find a very noticable rhizocephalan externa on the abdomen of several of the shrimp. Click here to learn more about the bizarre life of rhizocephalans.
Because of the hard calcareous plates that protect the body of most cirripeds, there was a good likelihood they would leave fossils. Likewise there is a good fossil record for the cirripeds stretching back as far as the middle Cambrian.
Introduction to Copepoda

Copepod, Calanus hyperboreus, courtesy of Uwe Kils.
Some would argue that the Copepoda is one of the most important groups of the crustaceans. They are found in abundance and constitute an important source of protein in the oceans. Their fecal pellets also accumulate onto the ocean floor and greatly accelerate the flow of nutrients and minerals from the ocean surface to the ocean floor.
Many copepods are parasites. However, several groups are free-swimming and are food to many fish and the whales of the northern hemisphere. You can read more about the different groups of copepods at the systematics page.
Often many copepod species occur in the same area. This observation raises questions about the resource partitioning of those copepods. Click life history to read more about copepod ecology and behavior.
| Copepod bodies are divided into two regions, the forward region is the prosome and the back region the urosome. These regions are separated by the major articulation or flexing point in the body. Different copepod groups have different numbers of segments in the prosome, so no generalization can be made. Calanoid copepods have enlarge first antennae which they often use for swimming. Calanoids can also be distinguished by a broad prosomes and narrow urosome whereas harpactacoids prosome and urosome are similar widths. In both groups all appendages are found on the prosomal segments except for the spiney appendages on the last body segment called the caudal ramus. |
|

Introduction to the Malacostraca
Crabs, shrimp, beach hoppers, krill, pill bugs, and mantis shrimp
There are more than 20,000 species of Malacostracans described to date. Members of Malacostraca generally have a total of 19-20 segments; 5 segments make up the cephalon or head, 8 segments compose the thorax, and 6 or 7 segments make up the abdomen. It is difficult to generalize about Malacostracans because the group is so large and diverse. For example, some have a carapace and some do not.
Malacostraca can be divided into two groups, the Phyllocarida, and the Eumalacostraca. Phyllocarida contains the oldest crustacean known and includes only one living group. They are also the only Malacostracans with phyllopodus (leaf-like) appendages. The Eumalacostraca consists of all Malacostracan groups other than the Phyllocarida. Eumalacostracans generally possess a well-developed carapace and a long, muscular abdomen. Eumalacostraca is the group that contains most of the animals the general public recognize as crustaceans, such as shrimp, crabs, lobsters. Choose from the cladogram below to begin exploring the Malacostraca.
Crustaceamorpha:Part II
Malacostraca


Introduction to Stomatopoda
Mantis Shrimp


Stomatopods are commonly called "mantis shrimp" for an obvious reason. Despite their importance to reef communities, little is known about them. We do know that they have a rather long fossil record that dates back 400 million years. Such a long fossil record helps researchers understand the evolution of some the more intriguing features of this group—their mantis-like arms and the eyes. Stomatopod eyes, which allow them to see polarized light, are some of the most complex among animals.
| Second thoracopod of a spearer (top) and and a smasher (bottom). Image provided with permission from Roy Caldwell, U.C. Berkeley. |
|

The most distinctive feature of a stomatopod is the raptorial second thoracic appendages. These are massive appendages that come in two basic types, smashers or spearers. Smashers have a thickened, hard pad on what could be loosely compared to their elbow. They use this pad to strike at passing prey or rival stomatopods. They are capable of crushing mollusk shells and instantly killing prey with a blow of this structure. This fact makes some of the larger species difficult to keep in aquaria since they are capable of breaking the glass with one quick blow. The spearers have tough spines instead of a hard pad on the second thoracopod and use it to spear their prey. The force and speed generated by the second appendages is astounding.
A stomatopod's long abdomen has little musculature compared to a lobster. The abdomen supports the flap-like pleopods which hold the gills and are also used for swimming. The tailfan, made up of the telson and uropods, is often elaborate with spines or other armor and vivid coloration. The tailfan is used to hold the abdomen and gills above the bottom, to block the burrow entrance, and to signal other stomatopods.
Visit the UCMP special exhibit Secrets of the Stomatopod: An Underwater Research Adventure to learn more about some of the unique features of stomatopods (including the eyes and raptorial appendages), and about a 2001 research expedition to study stomatopods in their own habitat.
Introduction to Branchiopoda
Sea monkeys, tadpole shrimp, water fleas, and clam shrimp


Left: A mirrored right lateral view of Daphnia magna, courtesy of www.splash.org
Right: A right lateral view of a fairy shrimp, courtesy of U.S. Fish and Wildlife Service
The Branchiopoda consist of four living groups: the Anostraca ("fairy shrimp" or "sea monkeys"), Notostraca ("tadpole shrimp"), Cladocera ("water fleas"), and Conchostraca ("clam shrimp'). There are also two fossil groups that belong to the Branchiopoda, but it is unresolved whether they are subgroups within the living groups or evolved separately. Here the fossil groups will be discussed within the living groups to which they are most similar.
With almost 800 described species, it is difficult to generalize about the branchipods. Most live in fresh or brackish (slightly salty) water and a few are found in marine habitats. Many are found exclusively in temporary ponds, where their eggs survive long periods of drought. You often find packages of dried eggs for sale in pet and toy stores. When the eggs are placed in water with an airstone, they hatch in just a couple days and you can watch them grow and swim. They are often used as food for aquarium animals.
It is even more difficult to generalize about the body form of branchiopods. The thorax and abdomen are fused or indistinguishable in most but the Anostraca. Their appendages are generally phyllopodus (leaf-like), although some groups have more appendages than others. They have a combination of paired compound and/or a single simple median eyes. Many are capable of parthenogenesis (growing from unfertilized eggs) but several use other reproductive strategies, ranging from releasing eggs when the adult molts to encapsulating the eggs in a modified molt.
Branchiopods feed in many ways. Some suspension feed and either remove organic particles from the water as they swim or stir up sediments to suspend organic particles that have settled and then remove them from the water. Others scrape organic matter from sand grains and rock. Still others actively prey on other small animals.
The most familiar member of the Anostraca are Artemia. They have elongate bodies compared to most other living branchiopods, with leaf-like appendages that they use to swim belly up. One centimeter is the average size for adult anostracans but some species will reach as much as 10 cm. Anostracans completely lack a carapace. The extinct group Lipostraca is often joined with the Anostraca. The Lipostraca consist of one species, Lepidocaris rhyniensis Scourfield, 1926, found in a very unusual fossil formation, the middle Devonian Rhynie Chert of
The Notostraca are called "tadpole shrimp" because of their broad, plate-like carapace and narrow, elongate thorax/abdomen. When populations are very large, Triops, a well known notostracan, can cause considerable damage to rice paddies by burrowing into the sediment and inadvertantly dislodging the young shoots. Notostraca has only 9 species, considerably fewer than any other branchiopod groups. The species are rather similar to each other and practically indistinguishable from extinct fossil forms. A fossil group often paired with the Notostraca are the Kazacharthra, found only in Jurassic rocks of Kazachstan, a country that neighbors Afganistan. They differ from notostracans by the shape of their carapace, which has elaborate spines on the margin giving them a rather fierce appearance.
Cladocerans are called "water fleas" and are typically quite small ranging from one half millimeter to three millimeters (however, oone species reaches up to 18 mm). The best known member of the Cladocera is Daphnia. The carapace of cladocerans covers the throrax/abdomen but not the head, and is folded down the midline giving them a bivalved appearance. Cladocerans dominate the plankton found in freshwater habitats. The cladocerans are mostly benthic but those that do swim, do so with their antennae instead of their thoracic appendages. The few cladoceran groups that occur in marine habitats are very different, having huge eyes and instead of being filter feeders like most of their freshwater relatives are active predators.
The Conchostraca or "clam shrimps" differ from the Cladocera in three primary ways: 1) They have appendages along the entire thorax/abdomen whereas the appendages are found only on the anterior segments of cladocerans. 2) Their carapace is bivalved with a hinge that allows opening and closing of the two halves. The cladocerans' carapace, on the other hand, is a single piece that wraps around the animal to cover the animal. 3) The conchostacan body is completely enclosed in the carapace but the head (cephalon) of cladocerans is not covered by their carapace. Conchostracans are found exclusively in freshwater.
Introduction to Stomatopoda
Mantis Shrimp


Stomatopods are commonly called "mantis shrimp" for an obvious reason. Despite their importance to reef communities, little is known about them. We do know that they have a rather long fossil record that dates back 400 million years. Such a long fossil record helps researchers understand the evolution of some the more intriguing features of this group—their mantis-like arms and the eyes. Stomatopod eyes, which allow them to see polarized light, are some of the most complex among animals.
Introduction to Remipedia
Dorsal view of a remipede from the
Photo courtesy of Thomas Iliffe (Texas A&M).
 The Remipedia are a small crustacean group. Its members consist of a handful of living species and a single fossil. Living representatives of this group, called the Nectiopoda, were not found until the early 1980's and there are still only seven species described to date from the
The Remipedia are a small crustacean group. Its members consist of a handful of living species and a single fossil. Living representatives of this group, called the Nectiopoda, were not found until the early 1980's and there are still only seven species described to date from the
Remipede bodies are divided into a cephalon (head) and elongate trunk with up to 32 similarly shaped segments. All known living remipedes except Enantiopoda lack eyes. The appendages on the trunk segments are paddle-like and directed out to the side. This appendage orientation is unusual in crustaceans, and it makes them superficially similar to polychaete worms (Annelida). Remipedes swim on their backs similar to the Anostraca (fairy shrimp).
There remains much to learn about the biology of these unusual animals.
Introduction to the Cephalocarida
The nine described species of cephalocarids are benthic (on the sea floor) marine animals. They are found from the intertidal zone to depths of 1500 m. Cephalocarids feed on detritus (dead organic material) and are found in all kinds of sediment, from soft, silty mud to clean course sand.
The tiny (2-3.7 mm long—that's about the size of a sesame seed) body of a cephalocarid is elongate and fairly simple compared to most crustaceans. Most specialists think of them as rather primitive even though they have not yet been found in the fossil record. They consist of a head, a thorax with 8 segments that possess biramous, paddle-like appendages, an abdomen that consists of eleven segments with no appendages, and a telson. They lack eyes.
Introduction to the Uniramia - Insects and their Relatives
Beetles and millipedes and flies and centipedes and....

The largest major group of arthropods is a clade that includes insects, millipedes, centipedes, and their relatives. This group, the Uniramia, was formerly defined to include the Onychophora, which are now considered a separate clade. It is restricted here to include only "true" arthropods with exoskeletons and jointed appendages.
Uniramians have strictly uniramous appendages; that is, their legs have only one branch. Most are terrestrial, but some are aquatic for part or all of their life cycles. They make up by far the most common and diverse major clade of arthropods, and in fact make up over three-fourths of all known animal species on the planet -- and probably an even greater proportion of the total number of species, known and unknown. As the paleontologist J. Kukalová-Peck once put it: To a first approximation, every animal is an insect. Think about that the next time you call Orkin Pest Control.
Uniramia:Part I

It isn't easy to condense the diversity of over three-fourths of the world's animals into a few pages, but we went ahead and did it. This phylogeny has been generally accepted for the past fifteen years, and so we use it here.
Where are the insects on this page? Insects are distinguished by having a body divided into head, thorax, and abdomen, with 11 segments on the abdomen, and three pairs of appendages on the thorax. Thus the Myriapoda, the centipedes, millipedes, and their kin, are not insects; they have many more appendages, and their bodies are not divided into thorax and abdomen.
Neither are the Parainsecta (springtails and proturans) or Diplura (diplurans) true insects, for their numbers of abdominal segments vary. These two clades, together with the true insects, form the clade Hexapoda ("six-legged"). The Insecta include all clades clockwise from the Archaeognatha (jumping bristletails); thus it also includes the Thysanura (silverfish or firebrats), Ephemeroptera (mayflies), Odonatoida (dragonflies and damselflies), and the many insect orders in the Neoptera, as well as other groups known only from fossils.
Parainsecta
Springtails and Proturans

The Parainsecta (which may be a paraphyletic group) include a number of small, wingless uniramians, divided into two classes: the Collembola (springtails) depicted in the upper part of this drawing, and the Protura at the lower left and center. Unlike true insects, whose abdomen is always composed of 11 segments, collembolans have only six abdominal segments, and proturans have 9 to 12. The Collembola possess a jumping organ that fold back under the abdomen, the furcula - from which they have received their common name of "springtails." Members of both classes are small and inconspicuous, but may be quite common in leaf litter, in soil, or on moss.
Collembolans have the honor of being the oldest known hexapods (six-legged uniramians) in the fossil record, predating the oldest true insects. The Rhynie Chert, from near
Orthoptera
Crickets and Grasshoppers



Click on either picture to receive a full-sized version!
The name Orthoptera is sometimes used for all the insects in the "orthopteroid" assemblage, including roaches, earwigs, mantises, and many others. However, it is more usual to restrict the Orthoptera (Greek for "straight-wing") to the crickets, grasshoppers, katydids, and their kin. These insects are instantly recognizable by their long hind legs, which are modified for jumping. Most orthopterans can generate noise by rubbing special organs together on their legs or on their wings, a habit known as stridulation. This photograph, taken with the UCMP Environmental Scanning Electron Microscope, shows the "file" -- a ridged vein on the wings of a mole cricket in the genus Oecantha. Mole crickets rub this file over another specialized structure on the other wing, the "scraper," to produce sound. Special membranes on the legs or on the abdomen allow orthopterans to hear these sounds, which are used in courtship.
The mouthparts of orthopterans are modified for chewing plant material; in fact, orthopterans can be serious agricultural pests in certain areas.
The oldest fossil orthopterans are Pennsylvanian in age; that is, about 300 million years old. The fossil grasshopper shown at the right of the page is much younger, only a few tens of thousands of years old; it is from the McKittrick Asphalt of southern California, a fossil-bearing tar deposit (very much like the famous La Brea tar pits in Los Angeles). Today, there are about 80,000 species of orthopterans. Some, like the swarming locusts, are serious pests and destroyers of vegetation. Others, we are told, are remarkably tasty and nutritious when properly prepared.
Introduction to the Myriapoda
They've got legs. . . they know how to use them. . .



Nearly 13,000 species of arthropod are classified in the Myriapoda, the "many-legged ones." All myriapods are terrestrial forms. Like insects and other uniramian arthropods, myriapods have appendages with only one branch, or ramus. Myriapods can have anywhere from fewer than ten to nearly 200 pairs of appendages; they range in size from nearly microscopic to 30 cm in length. Most myriapods live in humid environments, and can be found in soils, in leaf litters, or under stones and wood. Many species possess repugnatorial glands, specialized glands that secrete foul-tasing compounds and thus function in defense.
There are four groups of myriapods; how they are related to each other is not yet well understood. Two of them, the Symphyla and Pauropoda, consist of tiny arthropods living in leaf litter and soil; both superficially resemble centipedes. The Chilopoda includes the true centipedes, like the one shown at the top left of this page. Chilopods have only one pair of legs per body segment. They are predators; the first pair of appendages on the trunk are modified into a pair of claws with poison glands, which centipedes use to capture prey (usually other arthropods). The bite of large centipedes can cause humans some pain and discomfort, although there are no authenticated cases of human fatalities from centipede bites.
Myriapods in the Diplopoda ("double legs") include the millipedes, like the one pictured at the top right of this page. Millipede segments are formed in early development by fusion of two adjacent embryonic segments; thus, each adult segment of a millipede bears two pairs of legs. Unlike the predatory Chilopoda, most millipedes feed on decaying vegetation, although some are carnivorous.
Because most myriapods have a thin, light cuticle and live in environments where fossilization is unlikely, the fossil record of myriapods is quite sparse. Enough remains, however, to show that the group is a very old one indeed. The oldest fossil uniramians are myriapod-like marine organisms from the Cambrian. Some fossil burrows from the Ordovician have been claimed as myriapod burrows; this speculation is hard to test, but if it is correct, then myriapods might have been living on land as early as 400 million years ago. The oldest definite body fossils of myriapods come from the Late Silurian, though the oldest centipedes come from the
Introduction to the Odonata
Dragonflies and Damselflies
The order Odonata ("toothed ones") includes some of the most ancient and beautiful insects that ever roamed Earth, as well as some of the largest flying invertebrates ever to have lived. Odonata consists of three groups: Anisoptera (which includes dragonflies), Zygoptera (which includes damselflies), and Anisozygoptera (a relict group represented by only two living species. This order is very diverse with about 5000 species, and its members are easy to observe.
|
|
|


Adult Odonata : At left, an adult dragonfly (Anisoptera). Notice the relatively stout body, and the wings extending to each side while the dragonfly is perched. At right, an adult damselfly (Zygoptera). Damselfly bodies are slender by comparison, and wings are held above the body while at rest.
Both dragonflies and damselflies belong to the Odonata, which is a subgroup of insects, which in turn is a group of uniramian arthropods. Many characteristics distinguish Odonata from other groups of insects -- minute antennae, extremely large eyes (filling most of the head), two pairs of transparent membranous wings with many small veins, a long slender abdomen, an aquatic larval stage (nymph) with posterior tracheal gills, and a prehensile labium (extendible jaws underneath the head). Among living Odonata, there are twenty-five families, mostly dragonflies and damselflies. Of all their characteristics, the easiest way to tell a dragonfly or damselfly from other insects is by the size of the eyes and shape of the abdomen. If the eyes are very large in proportion to the head and the abdomen is long and thin, then it is almost sure to be in Odonata.
While both dragonflies and damselflies belong to the Odonata and share many common features, then are a number of noticeable differences as well. Even before hatching from the egg, differences in morphology of the egg distinguish dragonflies (Anisoptera) from damselflies (Zygoptera). Dragonfly eggs are round and about 0.5 mm long, whereas damselfly eggs are cylindrical and longer, about 1 mm long. Similarly, the nymphs (larvae) of the two groups differ. A larval damselfly abdomen is longer and narrower with three fin-like gills projecting from the end. Dragonfly nymphs are shorter and bulkier, and the gills are located inside the abdomen. The dragonfly nymph expands and contracts its abdomen to move water over its gills, and can squeeze the water out rapidly for a short burst of underwater jet propulsion.
Most of a dragonfly's life is spent in the larval stage where it molts from six to fifteen times. Depending on altitude and latitude, larval development varies from the common one or two years to as many as six years. At that time, the nymph crawls up out of the water and molts one last time, emerging from its old skin as an adult with functional wings. Unlike butterflies and beetles, dragonflies and damselflies do not have an intermediate pupal stage before becoming an adult. Because of this, Odonata are said to be hemimetabolous, or undergo an "incomplete" or "gradual" metamorphosis.
Larval Odonata : Dragonflies and damselflies begin their lives as nymphs, living underwater for a year of more. On the left, a damselfly nymph, recognizable by the three feathery gills extending from the tip of its abdomen. On the right, a dragonfly nymph, whose abdomen ends in three short spines and whose gills are held internally. The nymphs are not as brightly colored as the adults, but are well camouflaged predators who ambush their prey.
Both major suborders have large heads with very large compound eyes relative to the rest of their body. Each compound eye is composed of nearly 28,000 individual units (ommatidia), and together the eyes cover most of the head. More than 80% of their brain is devoted to analyzing visual information. By contrast, their antennae are tiny. Their mouths have been adapted for biting, making them efficient hunters. All Odonata have a prehensile labium, which can be extended forward from underneath the head faster than most prey can react, making their bite fatal to prey. The six legs are all located near the head and are seldom used for walking, but are more useful in catching prey and perching on vegetation to rest or lay eggs.
Both dragonflies and damselflies have two pairs of elongated membranous wings with a strong crossvein and many small veins that criss-cross in the wings, adding strength and flexibility to the wings. Both groups also have a characteristic nodus, or notch, in the front edge of each wing. In dragonflies, the rear wings have a broader base and are larger than the front pair. Damselflies, by contrast, have front and hind wings similar in shape, and as a result they fly slower than dragonflies do. Also, dragonflies do not have hinges enabling them to fold their wings together when resting, though damselflies do. This feature of the wings is the key morphological feature distinguishing adult dragonflies from damselflies.
 Dragonflies can fly forward at about 100 body-lengths per second, and backwards at about 3 body-lengths per second. They are also capable of hovering in the air for about a minute. Longer periods of stagnant flight would interfere with thermoregulation. The wings of male dragoinflies are relatively longer and narrower than females in large species. Adult wingspans measure from 17 millimeters (Agriocnemis) to 20 centimeters (Coerulatus). Most temperate zone species have wingspans of 5 to 8 centimeters and wings that are from two to twelve centimeters from front to back.
Dragonflies can fly forward at about 100 body-lengths per second, and backwards at about 3 body-lengths per second. They are also capable of hovering in the air for about a minute. Longer periods of stagnant flight would interfere with thermoregulation. The wings of male dragoinflies are relatively longer and narrower than females in large species. Adult wingspans measure from 17 millimeters (Agriocnemis) to 20 centimeters (Coerulatus). Most temperate zone species have wingspans of 5 to 8 centimeters and wings that are from two to twelve centimeters from front to back.
| |
Ancient wings : A fossil of Protolindenia wittei, a dragonfly with a wingspan of about 15 cm. This exquisitely preserved specimen comes from the Jurassic Solnhofen limestones of
The Odonata are known to be ancient insects. The oldest recognizable fossils of the group belong to the Protodonata, an ancestral group that is now extinct. The earliest fossils so far discovered come from Upper Carboniferous (Pennsylvanian) sediments in
Fossilized specimens of another group, the Protoanisoptera (family Meganeuridae), have been found in limestone at Elmo near
Though the Carboniferous specimens are the oldest fossils of this group found to date, they were not the first specimens to be discovered. The first Odonata fossils were found in sediments from the Lower Permian, over 250 million years old. These fossils are not huge monsters like the Carboniferous fossils, but belong to relatively small Protoanisopterans and Zygopterans (damselflies). The latter seem to have changed little in structure and appearance since then. However, it is currently a question of debate as to whether members of Protodonata and the earliest Odonata had aquatic larvae, as do all modern species, since no Paleozoic larvae fossils are known. Larvae do not exist as fossils before the Mesozoic. Some workers believe that Odonata adopted an aquatic larval stage during the Lower Permian, perhaps because their prey lived in aquatic habitats. In any event, several groups of Odonata existed by the Late Paleozoic, though only three members of this group survive today.
|
|
|
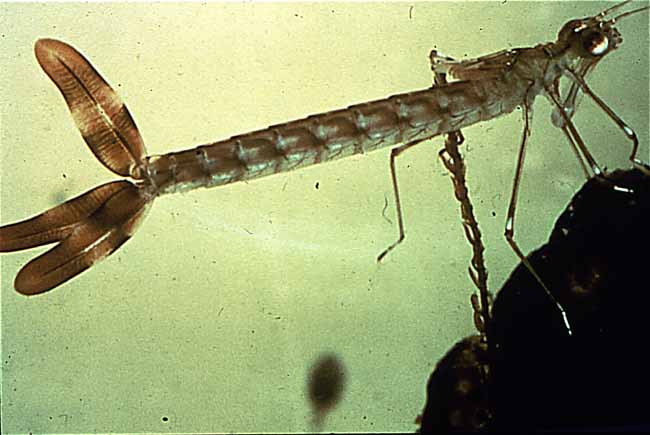

Generation gap : On the left, the nymph (larval form) of a damselfly. Notice the three large gills that the animal uses to "breathe" in its underwater environment. On the right, an adult dragonfly laying eggs. The transformation to the adult form sheds the gills in favor of two pairs of wings.
Dragonflies are generalists, that is, they eat whatever suitable prey is abundant. Oftentimes, they hunt in groups where large numbers of termites or ants are flying, or near swarms of mayflies, caddisflies, or gnats. According to most studies, the main diet of adult odonates consists of small insects, especially Diptera (flies). Maturing dragonfly larvae feed very intensively, as do females when developing their eggs. Studies show that food shortage may limit reproductive behavior. Dragonflies do not hunt in cold weather. Damselflies, however, are not as limited by temperature and have been observed hunting during cold spells. Males are territorial, sometimes patrolling for prey for hours at a time.
Though dragonflies are predators, they themselves must be wary of many predators. Birds, lizards, frogs, spiders, fish, water bugs, and even other large dragonflies have all been seen eating odonates. However, dragonflies have many adaptations enabling them to avoid predation. They have exceptional visual responses and truly agile flight.
Although many insects perform courtship, it is uncommon among dragonflies. Anisoptera copulate while in flight, the male lifting the female in the air. Zygoptera copulate while perched, sometimes flying to a new perch. The length of time required for copulation varies greatly. Aerial copulations may last mere seconds to one or two minutes. Perched copulations usually last from five to ten minutes. Intraspecific competition amongst males for females is fierce. It has even been discovered that in some species of Odonata, the males will remove all the sperm of rival males from a female's body before transferring his own sperm. These species are equipped with a "scoop" at the tip of the male's abdomen that is used for this purpose.
The distribution of various groups and species of Odonata is highly variable. Some genera and species are widespread while others are highly local in their distribution. Some families are restricted to cool streams or rivers, others to ponds or still clear waters, and some to marshy places. The presence of dragonflies and damselflies may be taken as an indication of good ecosystem quality. The greatest numbers of species are found at sites that offer a wide variety of microhabitats, though dragonflies tend to be much more sensitive to pollution than are damselflies. Many ecological factors affect the distribution of larvae. The acidity of the water, the amount and type of aquatic vegetation, the temperature, and whether the water is stationary or flowing all affect the distribution of Odonata larvae. Some species can tolerate a broad range of conditions while others are very sensitive to their environment.
Mayflies
| Ephemeroptera is a group of 2,000 insect species commonly known as mayflies. They are considered to be part of the clade Uniramia which includes silverfish and dragonflies, among others. Ephemeroptera and Odonata are the only extant orders of winged insects in the infraclass Paleoptera. All other insects with wings are in the Neoptera, and are characterized by a wing articulation (joint) that allows them to fold their wings back over their abdomens at rest (Carpenter, 1992). Ephemeroptera are aquatic insects that often go through many nymph stages (living in water) and two flying stages (the subimago and the imago). They are the only insects to have two flying stages, and can be recognized by their three caudal filaments (tails) at the tip of the abdomen, and a single claw on each leg. This differentiates them from the closely related stoneflies which have two tarsal claws. The flying stages are characterized by relatively large forewings, which are usually kept upright, and reduced or nonexistent hind wings. |
|

Mayfly Morphology
The first stage of the life of a mayfly is the nymph (larva), which not only looks very different from the adult, but lives in the water. When the nymphs hatch from the eggs, they are less than 1 mm long. They have no gills at first, and their body shape varies according to habitat. For example, those that burrow (such as Ephemera) have more cylindrical bodies, whereas those that slide under rocks (such as Heptagenia) are flatter. Those in the genus Caenis crawl on mossy stones and vegetation, so they have short bodies with squat legs. Ephemeroptera nymphs may grow to anywhere from 4 mm to 3 cm long. They are generally camouflaged against their background. The number of molts a nymph goes through on its way to becoming an adult does not depend on its nutrition, but the increase in size that comes with each molt does.
In older nymphs, gills are found in pairs on each segment of the abdomen (see pictures below). The gills extend from the sides of the body and are oval-shaped. These gills beat to control the flow of water through the body, which also controls the amount of oxygen and salt that flows through the body. Nymphs in still waters generally have larger gills, and those in running water have smaller gills; this allows the nymphs of each habitat to get their optimum flow of water. Not only do the gills function in uptake of water, salt, and oxygen, but they also send water off at right angles to the body. This is used to mislead predators. If the water simply flowed out the back of the nymph's body, predators would know that the nymph was sitting at the beginning of the stream. However, since they send water away from their bodies at several points, the nymphs are not as easy to track.
Some signs of sex can be seen in the last few stages of the nymph, even before it becomes an adult. At this stage, male nymphs have the beginnings of clasping organs on the lower portions of their abdomen, with which they hold the female during copulation. In some species, the males have divided eyes that are two colors. The upper portion is for seeing movement, and the lower portion is specialized for seeing details. The females have smaller eyes and oviducts in the lower abdomen.
When it comes time for the last nymph stage to molt into a subimago (the first flying stage), the guts empty out and the mid-gut section fills with air. Often, many nymphs will then simultaneously let go of their hold on their anchor in the water and float up to the top. Once they reach the air, the cuticle splits open on the thorax and the wings come out. This is the time of greatest vulnerability in their lives as they float on the water before they are strong enough to fly. The subimago has short hairs on the wings and on the body; the wings are dull and pigmented. Once it gains some strength, it flies from the water to some form of shelter such as a tree, long grass, or the underside of a bridge and molts again within 24 to 48 hours. Thisadditional molt allows the legs and tails of the insect to grow more. Longer tails give more stability in flight, and longer legs make it easier for the male to grasp the female in mating.
|
|
|


The imago (the final adult stage) has shiny, hairless wings. The longer legs and tails allow for more rapid flight. The corrugation of the wings protects them by making them more flexible and therefore less vulnerable to wind damage. The imago mates and dies within a few hours to a day. (Harker, 1989) This short adult life is what gives the order its name from the Greek ephemeros meaning "lasting but a day."
Mayfly nymphs : At left above, is the nymph of a Baetid mayfly, and at right is a Heptageniid nymph. Notice the Baetid has a slender, cylindrical body and small gills on its abdomen. The small gills indicate that it lives in moving water, and the shape of its body makes it well-suited for swimming against the current. The Heptageniid has a broad flat body, better suited for life clinging to the bottom of the stream to avoid being carried away by the current. Notice also that both nymphs lack the wings of the adult stage, and both have the three caudal filaments (tails) characteristic of mayflies. (Click on either of the pictures above for a larger image).
Mayfly Ecology
Mayfly eggs are eaten by snails and by caddisfly larvae. The nymphs may be eaten by fish, frogs, birds, flies, or water beetles. The subimagos are eaten by fish, birds, dragonflies, water beetles, or other predatory insects. For their own nutrition, mayfly nymphs move over stones and weeds to graze off bacteria. They may collect from sediments or feed on detritus. Most mayflies are collectors and scrapers (Edmunds et al, 1976), and most of the consumed bacterial cells pass through their bodies without being used. They are opportunistic generalists, meaning that they eat what they can, when they can. Nymphs have some symbiotic relationships with chironomids that may be commensal. The chironomid larvae attach to the cuticle of the mayfly and eat the unicellular organisms that live there. More harmful to mayflies are nematode parasites, who use nymphs as their hosts by eating their muscles from the inside.
Ephemeroptera nymphs are usually microhabitat specialists. Each species survives best on a specific substrate at a certain depth under water with a certain amount of wave action. For example, Rithrogena generally live in medium to large trout streams. Ephemeridae burrow into soft areas where flow is slower, or in areas of lakes and rivers where deposits occur; the particular substrate and burrow depends on the genus. The primitive habitat of schistonate mayflies is still water even though most extant mayflies live in running water (McCafferty, 1990). In some areas, succession occurs by different species. For example, in
Mating occurs in a swarm, and at these times there may be such dense clouds of mayflies in the air near streams that driving becomes impossible. Because their wings are so fragile, the imagos need calm weather in order to mate. Males usually swarm very near the water, though swarm formations vary from species to species. Within the swarm, the insects are always changing positions. When a female enters the swarm, males try to mate with her. Some float to the ground while mating, and others continue flying. Once a male has successfully mated, he will guard the female to make sure that no other male mates with her. The female then flies to water to lay her eggs. She dips into the water while flying and releases a few eggs each time. The eggs sink to the bottom and their surface changes. Some become covered with a sticky substance and some have adhesive disks. Some species are parthenogenic, meaning that they do not need sperm to produce fertile eggs (Harker, 1989). The time it takes for emergence into the subimago form varies depending on temperature -- the milder the temperature, the earlier the emergence (Edmunds et al, 1976). Many species have synchronized emergence of subimagos. In these cases, the emergence occurs at a specific time of day under certain weather conditions (Harker, 1989).
Mayfly Fossil Record
The first recorded mayfly nymphs are from the Late Carboniferous (Fenton, 1989). In most places, mayflies are represented primarily by larval fossils (Sinitshenkova, 1990). Presumably, this is because of the very short lifespan of the adult stage. The siphlonurid form, which is shrimp-like with short caudal filaments that have lateral hair fringes, is the primitive form (Riek, 1970). However, it is difficult to classify early fossils in the correct order. The fossil Triplosoba pulchella is the only insect from the Carboniferous that is consistently placed in the order Ephemeroptera (McCafferty, 1990).
In recent years, certain fossils found in
Early mayfly adults differ from the Permian also differ from their living descendants. Fossil imagos of Protereisma from
Currently, the Ephemeroptera are classified in several different ways, depending on who does the cladistic analysis. According to Riek, this order has six superfamilies with 14 families. There are also two extinct superfamilies, Proterismatoidea and Mesephemeroidea, which are the Permian insects that appear to be mayflies or their precursors. During the lower Cretaceous, there was extinction and emigration of many mayflies in
Uniramia:Part 2
The Neoptera

The Neoptera contains most of the diversity of the insects -- that is to say, most of the diversity of the animals on this planet. Three of the groups shown on this page, the Coleoptera (beetles), Lepidoptera (butterflies and moths), and Hymenoptera (ants, bees, and wasps), together account for about half of all known animal species -- and new species of these and other insect groups are being discovered almost every day.
Don't be put off by the long Latin names on this cladogram. Most of them refer to very familiar groups of insects:
- Plecoptera -- stoneflies
- "Orthoptera" -- roaches, mantises, crickets, and many others
- Phthiraptera -- lice
- Psocoptera -- booklice
- Hemiptera -- true bugs, aphids, cicadas, and others
- Thysanoptera -- thrips
- Strepsiptera -- twisted-wing parasites
- Coleoptera -- beetles
- Neuropterida -- lacewings, ant lions, dobsonflies
- Hymenoptera -- ants, wasps, and bees
- Lepidoptera -- butterflies and moths
- Trichoptera -- caddisflies
- Diptera -- true flies
- Siphonaptera -- fleas
- Mecoptera -- scorpionflies
Hymenoptera
Bees, ants, and wasps



Hymenopterans, the "membrane-winged" insects, include bees, ants, and a large number of other insect taxa collectively referred to as wasps. The Hymenoptera include famous examples of social insects, such as honeybees and true ants; these insects have developed regimented social systems in which members are divided into worker, drone, and queen castes. Such social hymenoptera may live together in nests or hives of many thousands of individuals, all descended form a single queen. Not all hymenoptera are social, however; many live a solitary life, coming together only for a brief mating.
As is obvious from the picture above, hymenopterans include important pollinators of flowering plants. Flowers pollinated by bees are typically yellow or blue and often have patterns visible only under ultraviolet light, which bees can see. Many bee-pollinated flowers are bilaterally symmetrical and produce abundant nectar, such as the orchids, some species of which depend on a single species of bee for pollination. Other plants may be pollinated by ants, or may rely on ants living within them to keep predators away. Many of these plants produce large quantities of nectar, or produce other fluids for the ants.
Less well-known, but equally spectacular, are the various families of parasitoid wasps; parasitoids lay eggs in living hosts, often other insects, which hatch into larvae that feed on the host's tissues before emerging. Such treatment typically kills the host; parasitoids are attracting interest as natural controls on insect pests. Other wasps parasitize the eggs or larvae of other insects, such as the wasp below, which is shown emerging from the egg of a hemipteran that was parasitized.
Hymenopterans have left behind some spectacular fossils. The best-preserved are those found in amber, which is actually fossilized sap from ancient trees. In these fossils, detail of legs, antennae, wings, and even small body hairs are preserved.
Phthiraptera
Lice

The Phthiraptera are the lice; this one, the human head louse Pediculus capitis, was drawn in the seventeenth century by the early microscopist and scientist Robert Hooke. Lice are insects that have secondarily lost their wings and become ectoparasites -- external parasites -- on birds and mammals. Sucking lice (Anoplura), such as the human body louse figured above, have modified mouthparts for piercing and sucking. Others, traditionally grouped in the Mallophaga, are referred to as chewing lice, because their mouthparts are modified for chewing. Phthirapterans show extreme host specialization; in fact, many bird and mammal species can be identified by nothing more than the lice species that live on them, for these lice species would be found nowhere else.
Lice have almost no fossil record; the only ones known have been recovered from frozen mammals in Siberian permafrost, and are about 10,000 years old. Since the Phthiraptera as we know them today are restricted to birds and mammals, lice probably did not come into existence before the Jurassic -- although ancestral Phthiraptera might have fed on the ancestors of birds (dinosaurs!) and of mammals in the Triassic.
Siphonaptera
Fleas

Ctenocephalides felis, the cat flea shown here, is a typical member of the Siphonaptera ("sucking-wingless"), the fleas. Although known to be related to the true flies (the Diptera) by details of internal structure, the Siphonaptera are highly modified for a parasitic existence; they have no wings and no compound eyes, the legs are modified for jumping, the antennae are very short and recessed in grooves on the head, and the body is highly flattened laterally. Most are parasites on mammals; a few live on birds. Adult fleas feed on blood, while their larvae feed on organic debris.
Fleas are extremely rare as fossils; their small size and specialized habitat makes them highly unlikely candidates for fossilization. Two species have been found in amber from the Baltic region (late Eocene-Oligocene); living members of the family of fleas to which these fossils belong (Hystrichopsyllidae) are mostly parasitic on insectivores (moles and shrews). A few Cretaceous fossils have been assigned to the Siphonaptera (e.g. Riek, 1970), but some of these fossils are questionable (Carpenter, 1992). Since the Siphonaptera are mostly parasitic on mammals, they presumably evolved at about the time the mammals were evolving, in the Jurassic and Cretaceous.
Coleoptera
Beetles

It has been estimated that half of all animal species alive today are beetles; using a conservative estimate of the number of animal species, there would be at least three million beetle species on the Earth. The Coleoptera also includes some of the largest insects, like the living Hercules beetle, Dynastes hercules, shown here.
Coleoptera means "sheathed wing;" beetles have two pairs of wings, but the first pair has been enlarged and thickened into a pair of hard sheaths, or elytra, that cover the delicate hind wings. Because the elytra are fairly hard structures, beetles have a better fossil record than many other insect groups do; the oldest fossil beetles are Permian. Below is a much later fossil beetle, from the Pleistocene-age McKittrick Asphalt of southern

Because beetle elytra may be common fossils in Pleistocene deposits, they are often important sources of data on Pleistocene environments.
Thysanura
Silverfish and firebrats

The Thysanura are a group of small wingless insects. While most species spend their time in decaying material, leaf litter and the like, a number of them have found urban living preferrable. Silverfish are commonly found in older buildings and any place they can live undisturbed on old newspapers and books. Firebrats, however, get their name from their preference for warm places, such as near ovens and heaters.
Thysanoptera
Thrips

Thrips are small insects, typically only a few millimeters long. The wings, fringed with setae, small hairs which are distinctive for the group and give it its name, which means "fringe-wing." Some thrips are predaceous, but many feed by sucking plant juices, and may be agricultural pests.
Most fossil thrips have been found in amber, where they are fairly common. The oldest fossil thrips are Permian.
Lepidoptera
Butterflies and Moths
 Butterflies and moths make up the Lepidoptera. The name means "scale wing," and lepidopteran wings are covered with microscopic scales, which are iridescent and brightly colored in the case of this
Butterflies and moths make up the Lepidoptera. The name means "scale wing," and lepidopteran wings are covered with microscopic scales, which are iridescent and brightly colored in the case of this
The first definite lepidopteran fossils appear in the Lower Cretaceous, though lepidopterans are not common as fossils. Their evolution has been closely tied with the evolution of flowering plants, which also appeared and began to diversify in the Cretaceous. Among the butterflies, both adult and caterpillar stages feed on flowering plants, and the adults are important pollinators of several flowering plant families.
"Orthopteroid" Insects
A number of major clades of insects have at one time or another been grouped in a loose assemblage of taxa, sometimes called the Orthoptera or Orthopteroida (meaning "straight-wing" in Greek). While it seems likely that these "orthopteroid" groups are fairly closely related to each other, exactly how they are related is not fully understood, although new molecular studies are providing some important information (e.g. Kambhampati, 1995). Since we are not sure whether the "orthopteroids" are monophyletic or paraphyletic, we have chosen simply to list the major groups. Eventually we will provide pages for these taxa.
- Blattaria -- roaches (pictured above)
- Isoptera -- termites
- Mantoidea -- mantises
The above three groups are sometimes classified together as the Dictyoptera, a grouping supported by molecular phylogenetic analysis (Kambhampati, 1995).
- Phasmida -- stick insects
- Dermaptera -- earwigs
- Orthoptera -- true crickets, grasshoppers, katydids
- Grylloblattodea -- mole crickets
- Embioptera, or Embiidina -- web-spinners
- Zoraptera -- zorapterans
- Titanoptera -- extinct insect group (Triassic)
- Protorthoptera -- extinct insect group (Pennsylvanian-Triassic); probably including ancestors or relatives of ancestors of other orthopteroid taxa, thus paraphyletic.
Blattaria
 Roaches
Roaches
The Blattaria includes the roaches, from the six-inch tropical roaches of
Roaches arose in the Pennsylvanian period, about 280 million years ago, and have not changed much since. Compare the photograph of the living cockroach at the top of the page with this picture, depicting the 260 million-year-old, three-inch fossil roach Mylacris, and you'll see that roaches have not undergone any sweeping changes in their form in all that time, a phenomenon known to evolutionary biologists as stasis.
Fossil Record of Arthropoda
Soft-bodied relatives of the arthropods, as well as trace fossils that were made by some arthropod-like organisms, appear in the Vendian. However, arthropods underwent rapid evolution in the Cambrian Period; Cambrian localities like the world-famous Burgess Shale in

The above image is the head and mouthparts of a fossilized larva of the water bug Schistomerus, found silicified in Miocene deposits (about 15 million years old) near
Life History and Ecology of the Arthropoda
It is almost impossible to generalize about the ecology and life history of arthropods; they are simply too diverse. Arthropods first appeared in the sea; trilobites were all marine. Most crustaceans are also marine, but one crustacean group has succesfully invaded the land, the Isopoda, a.k.a. "pillbugs" or "roly-polies." The vast majority of living arthropods belong to groups that are specialized for a land-dwelling existence. One of them, the African tsetse fly, is shown here as a reminder of how successful some arthropods have been at the expense of other organisms.
Morphology of the Arthropada
Arthropods have an exoskeleton of a tough compound called chitin. This outer shell provides the structure against which arthropod muscles pull, reduces water loss, and protects them from environmental dangers. To move inside such a rigid armor, the chitin is laid down in plates, with joints between them. This gives the arthropods their name, meaning "jointed feet".
Some arthropods, like the centipedes, millipedes, and insects, have legs with a single branch (uniramous appendages). The rest historically have legs with two branches (biramous appendages). The outer branch is often a flattened gill, while the inner branch is often used for walking or modified for grasping, chewing, or reproduction. A number of arthropods that now apparently have uniramous appendages, such as spiders and scorpions, are descended from ancestors that had biramous appendages: appendages may be modified in such a way that one branch is lost or concealed. Arthropods also have a hemocoel, an open body cavity in which blood flows and bathes the tissues and organs. The dorsal tubular heart is perforated by pores.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
No comments:
Post a Comment